
Intolérance au lactose
Lactose, lactase et intolérance au lactose
La famine et les infections pourraient être à l'origine de la tolérance au lactose des anciens européens
Résumé tout public
Le lait et les produits laitiers constituent aujourd'hui une part importante de notre alimentation. Cependant il existe dans nos populations deux types d'individus : ceux qui digèrent le lait sans problème et ceux qui ne le digèrent pas ou mal. Cette bipartition tient au fait que le lait contient un sucre, le lactose, dont la digestion en glucose et galactose dans l'intestin grêle nécessite l'action d'une protéine (enzyme), la lactase.
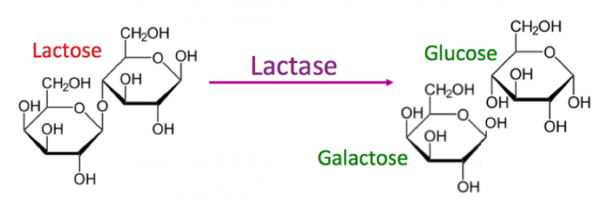
Lactose, lactase, glucose et galactose
Le lactose est le sucre principal présent dans le lait. Il n'est pas assimilable par l'intestin grêle. Son accumulation dans le colon provoque des troubles connus sous le terme "intolérance au lactose". La lactase est une enzyme qui scinde la molécule de lactose en glucose et galactose qui sont eux directement assimilables par l'intestin grêle.
Cette protéine est faiblement présente chez le fétus mais elle atteint un niveau élevé chez le jeune enfant (dont le lait est l'aliment principal) jusqu'au sevrage. Après le sevrage deux situations peuvent se présenter chez les adolescents et les adultes : ou bien la lactase reste à un niveau élevé, ou bien elle chute pour atteindre un niveau négligeable. Ceux chez qui le taux de lactase reste à un niveau élevé sont les porteurs du trait lactase persistante (LP) et peuvent digérer le lactose, alors que ceux chez qui ce taux est trop faible (porteurs du trait lactase non persistante, LNP) ne le digèrent pas. Ils sont dits "intolérants au lactose".

Les divers niveaux d'expression de la lactase chez l'homme
Présente à un faible niveau chez le fétus la lactase atteint un niveau élevé chez le jeune enfant jusqu'au sevrage. Après le sevrage la lactase peut rester à un niveau élevé ou chuter à un niveau négligeable. Les personnes porteuses des mutations génétiques -13910T et/ou -22018A (en haut à droite du schéma) gardent un taux de lactase élevé et peuvent digérer le lait. A l'inverse, celles qui portent les variants ancestraux (originaux) -13910C et -22018G (en bas à droite) ont un taux de lactase faible ou très faible et ne digèrent pas le lait. Elles sont dites "intolérantes au lactose".
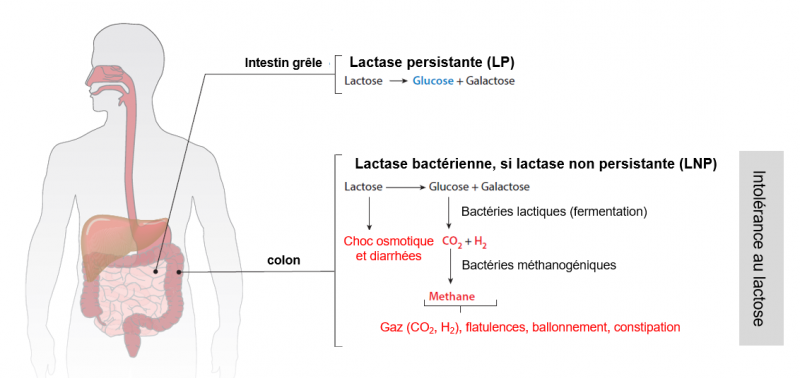
L'intolérance au lactose
Le lactose n'est pas assimilable par l'organisme. Pour ce faire, il doit être scindé en glucose et galactose par la lactase. Les individus qui expriment la lactase à un niveau élevé ne présentent donc aucun trouble après consommation de lait ou de produits laitiers. En revanche, chez les individus dont le niveau de lactase est faible, l'accumulation de lactose dans le colon peut entraîner un choc osmotique dont la conséquence est la survenue d'une diarrhée. De plus, la digestion du lactose est alors assurée par les bactéries lactiques qui provoquent une fermentation à l'origine de dégagement de CO2 et d'H2, et les bactéries méthanogéniques qui entraînent une libération de méthane, d'où les ballonnements, flatulences et dans certains cas constipation. L'ampleur et la gravité de ces symptômes peut fortement varier d'une personne à l'autre. Elle dépend de la quantité de lactose ingérée, du niveau d'expression de la lactase, du temps de transit intestinal, du microbiote intestinal, de la sensibilité individuelle et de facteurs psychologiques. Adapté de Ségurel et al. 2017.
Pourquoi cette bipartition lactase persistante versus lactase non persistante ?
En Europe et dans diverses populations d'Afrique, du Moyen-Orient et d'Asie du Sud, une vision consensuelle est que le trait de lactase persistante (LP) a été sélectionné par l'évolution au cours des 10 000 dernières années du fait de l'activité laitière mise en place par les premiers éleveurs durant le Néolithique.
Avant le Néolithique nos ancêtres étaient naturellement intolérants au lactose
Logique, le lait étant l'aliment spécifique du nouveau-né. Cependant, entre 10 000 et 5 000 ans avant J-C, au cours du Néolithique en Europe du nord notamment et d'autres régions du globe, plusieurs mutations sont apparues dans le génome humain (pour plus de détails voir "Pour aller plus loin"). Ces mutations ont eu pour conséquence le maintien de la production de lactase chez l'adulte. Les personnes affectées par ces mutations pouvaient donc consommer du lait sans en subir les troubles et leur trait de lactase persistante a été transmis à leurs descendants jusqu'à aujourd'hui.
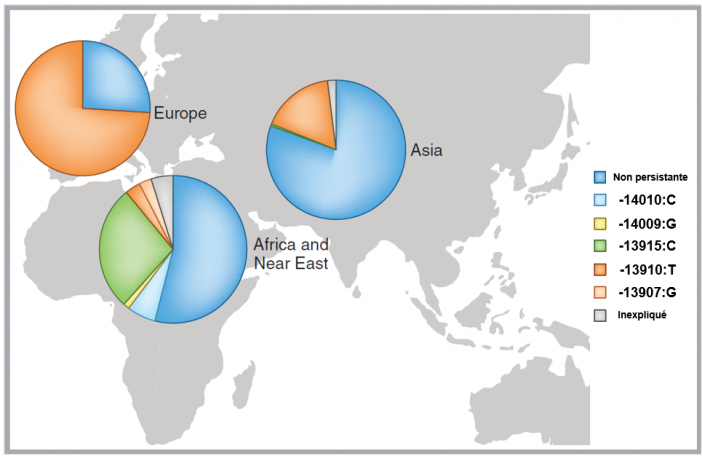
Distribution du trait de lactase persistante ou non persistante dans le monde aujourd'hui
Le trait de lactase persistante on non persistante a été évaluée par la fréquence des mutations génétiques fonctionnelles connues. Le segment bleu foncé montre la proportion approximative du trait lactase non persistante et les autres couleurs montrent la proportion du trait lactase persistante attribuable aux différentes mutations génétiques fonctionnelles décrites à ce jour : -13907:G, -13910:T, -13915:C, -14009:G, -14010:C décrites à droite de la figure. On voit clairement que le trait de lactase persistante est majeur en Europe, et essentiellement dû à la mutation -13910:T. On constate l'importance du trait lactase non persistante en Asie (>80%). Situation intermédiaire en Afrique du fait de la présence de différentes mutations. Adapté de Swallow et al. 2016.
Au niveau mondial, aujourd'hui, les personnes porteuses du trait lactase persistante représentent au total 30% de la population. Il y a donc eu une évolution favorable de ce trait pour les humains. Mais quel en a été le moteur ?
Deux hypothèses se sont affrontées :
- L'hypothèse "culture-historique" selon laquelle le trait de lactase persistante était rare chez nos lointains ancêtres jusqu'à l'avènement de la production laitière au début du Néolithique. Sa fréquence a ensuite augmenté rapidement sous l'effet de la sélection naturelle, du fait de l'avantage nutritionnel apporté par la consommation de lait et de produits laitiers.
- L'hypothèse de la " cause inverse ", selon laquelle la production laitière a été adoptée par des populations qui présentaient déjà des fréquences élevées du trait de lactase persistante. Pouvant naturellement consommer du lait sans dommage elles ont profité de cet avantage nutritionnel.
Cette deuxième hypothèse a toutefois été abandonnée lorsque les analyses de l'ADN ancien ont révélé l'absence de mutations conduisant au trait de lactase persistante chez les humains avant le Néolithique (Berger et al. PNAS2007).
Cependant, une grande incertitude demeure quant à aux facteurs qui ont contribué à cette évolution.
Un article récent publié en Juillet 2022 dans la revue Nature remet en cause l'hypothèse "culture-historique" et suggère que les famines et les maladies résultant de la proximité avec les animaux d'élevage (zoonoses) pourraient être à l'origine de la tolérance au lactose des anciens Européens.
En d'autres termes, pour nos ancêtres du Néolithique, le fait de pouvoir consommer du lait sans le moindre trouble, lors de conditions de stress nutritionnel sévère (famines) et/ou d'épidémies (zoonoses), aurait représenté un avantage suffisamment important pour favoriser l'évolution vers le trait de lactase persistante, jusqu'à nos jours.
Pour aller plus loin
La consommation de lait et de produits laitiers par des adultes peut être perçue comme une "hérésie biologique". En effet, la consommation par un adulte d'un aliment spécifique du nouveau-né a de quoi interroger. De fait, la nature a bien fait les choses : chez tous les mammifères, la production de lactase est stoppée dès la fin du sevrage. Chez l'homme cela se passe lorsque l'enfant atteint l'âge de 5 à 8 ans. Ce phénomène existe depuis la nuit des temps et bien avant la révolution néolithique. Cette hypolactasie (lactase non persistante ou LNP) qui est en fait la norme, interdit aux adultes qui possèdent ce trait la consommation de lait en les affligeant, en cas d'infraction d'un trouble appelé "intolérance au lactose" dont les symptômes sont des flatulences, des diarrhées, des crampes abdominales, pouvant aller jusqu'à des céphalées et des vomissements dans les cas les plus sévères. La gravité de ces symptômes dépend de la quantité de lactose ingérée, du niveau d'expression de la lactase, du temps de transit intestinal, du microbiote intestinal, de la sensibilité individuelle et de facteurs psychologiques. Ces symptômes apparaissent entre 30 minutes et deux heures après ingestion.
Cette intolérance au lactose ne doit pas être confondue avec les allergies au lait ou aux produits laitiers, qui se caractérisent par une réponse immunologique anormale pouvant évoluer vers une anaphylaxie grave mettant la vie en danger. Il existe trois formes principales de carence en lactase : congénitale, primaire et secondaire.
- Le déficit congénital en lactase (alactasie) est un trouble pédiatrique autosomique récessif rare associé à une absence d'expression de la lactase chez les nouveau-nés.
- Le déficit primaire en lactase (hypolactasie de l'adulte) est la condition naturelle résultant du déclin progressif et physiologique de l'activité enzymatique de la lactase qui se produit après le sevrage. Ce trait se transmet sur le mode autosomique dominant chez les homozygotes -13910:C/C et -22018:G/G.
- La carence en lactase secondaire, à l'inverse, peut être induite par une maladie ou une lésion de l'intestin grêle telle qu'une gastro-entérite, une maladie cœliaque, une maladie intestinale inflammatoire, une chimiothérapie ou un traitement antibiotique. Il est à noter que l'origine ethnique s'est avérée être un déterminant plus important de la susceptibilité à développer une déficience en lactase chez de nombreux patients atteints de maladies inflammatoires de l'intestin plutôt que les marqueurs de la maladie (en particulier pour les cas de colite ulcéreuse et de maladie de Crohn qui n'impliquent pas l'intestin grêle, d'obstruction distale ou de surcroissance bactérienne). Généralement, les étiologies non génétiques de l'intolérance au lactose peuvent être inversées si la cause peut être éliminée.
Cependant, il est clair qu'aujourd'hui chez environ un tiers des humains, l'expression de la lactase persiste tout au long de la vie, un phénotype connu sous le nom de "lactase persistante" (LP) qui leur permet de consommer du lait et des produits laitiers sans le moindre trouble.
Pourquoi cette bipartition lactase persistante versus lactase non persistante ?
Bien qu'aujourd'hui 95 % des Européens modernes, dans certaines régions (notamment les Iles Britanniques) soient porteurs du trait lactase persistante (LP), cela n'a pas toujours été le cas. Les données de séquence nucléotidique de l'ADN prélevé chez des individus "anciens" indiquent que la plupart, sinon la totalité, des personnes du Néolithique ancien étaient caractérisées par le trait de lactase non persistante (LNP, qui est encore aujourd'hui la norme) et que le trait de lactase persistante (LP) n'a atteint des fréquences significatives qu'aux âges du bronze et du fer.
Entre 10000 et 5000 ans avant J-C, au cours du Néolithique, plusieurs mutations sont apparues dans le génome humain dans la région régulatrice du gène de la lactase. Ces mutations ont eu pour conséquence le maintien de la production de lactase chez l'adulte par un passage du trait de lactase non persistante au trait de lactase persistante (LNP à LP). Les personnes affectées par ces mutations pouvaient donc consommer du lait sans en subir les troubles.
Mécanisme de régulation de l'expression du gène de la lactase (LCT)
En Europe, chez les personnes porteuses du trait lactase persistante, la mutation principale dans la région régulatrice du gène LCT est la mutation -13910:C>T. Il s'agit d'un SNP, "Single Nucleotide Polymorphism", en Français "polymorphisme d'un seul nucléotide". Cette notation C>T signifie que dans la forme ancestrale le nucléotide en position -13910 (par rapport au site d'initiation de la transcription du LCT) était un C et a été remplacé par un T chez les personnes porteuses de la mutation (Enattah et al. 2002; Lewinsky et al. 2005).
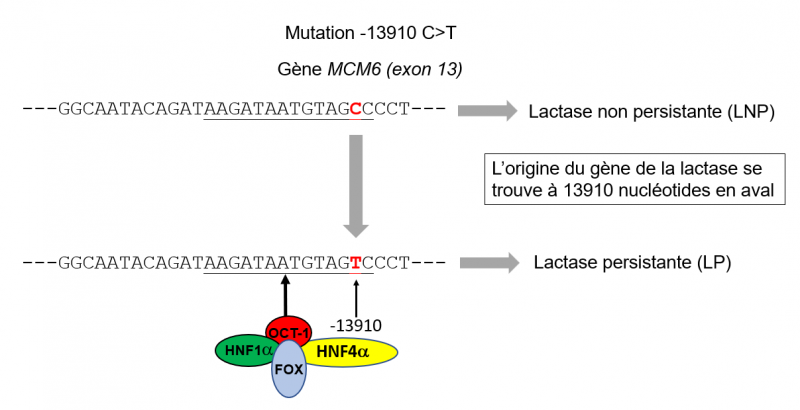
La mutation -13910:C>T
Une partie de la séquence de l'exon 13 du gène MCM6 est représentée ici. La séquence soulignée correspond au motif spécifiquement reconnu par le facteur transcriptionnel OCT-1. Ce facteur ne se lie apparemment pas sur la séquence ancestrale (séquence du haut de la figure) avec le nucléotide C en position -13910 (en rouge). Par contre il se lie sous forme d'un complexe avec trois autres facteurs (HNF1a, HNF4a et FOX) à cette séquence lorsqu'elle est mutée (le T en rouge remplace le C) et c'est cette liaison qui permet l'activation du gène de la lactase. Le site d'initiation de la transcription du gène de la lactase se trouve à 13910 nucléotides en aval (dans la direction 5'>3'). Adapté de Lewinsky et al. 2005.
Ce variant est l'un des exemples les plus probants de sélection naturelle sur le génome humain
Il est situé à 13910 nucléotides en amont du site d'initiation de la transcription du gène LCT dans l'exon 13 du gène MCM6. Ce site semble fonctionner en combinaison avec un autre site localisé dans l'exon 9 de MCM6 en position -22018:G>A. D'autres variants ont été découverts dans d'autres populations mais sont toujours situés dans la même région du génome comme le montre la table ci-dessous et produisent les mêmes effets en termes de lactase persistante (Ségurel et al. 2017).
Séquences ancestrales Mutations Aires Géographiques
-13910:C -13910:T Europe, Eurasie, Afrique du Nord, Moyen Orient
-13915:T -13915:G Moyen Orient
-13907:C -13907:G Afrique de l'Est (Ethiopie, Soudan)
-14009:T -14009:G Afrique de l'Est (Ethiopie, Soudan)
-14010:G -14010:C Afrique de l'Est (Kenya et Tanzanie) et Afrique du Sud
Le trait de lactase persistante se transmet sur le mode autosomique récessif, un seul allèle porteur d'une de ces mutations étant suffisant pour que le niveau de lactase soit maintenu à un niveau significatif.
A côté de ces 5 variants principaux, 18 autres mutations (rares pour la plupart avec une fréquence <5%) liées au trait de lactase persistante ont été trouvées : -22018:G>A (en combinaison avec la -13910:C>T), -14011:C>T, -13906:T>A, -13779:G>C, -13744:C>G, -13730:T>G, -13603:C>T, -13495:C>T, -13914:G>A, -14062:G>A, -14028:T>C, -13753:C>T, -13693:G>A, -13806:A>G, -13964:C>A, -13771:A>G, -14010 G>A, et -13926 A>C (Anguita-Ruiz et al. 2020). Ces mutations sont apparues principalement dans les populations en Afrique, au Moyent Orient et en Eurasie.
Des études de la régulation transcriptionnelle du gène LCT ont montré que la mutation -13910:C>T favorise fortement l'activité d'un facteur transcriptionnel, la protéine OCT-1. La figure ci-dessous est un schéma très simplifié de la région régulatrice d'un gène.
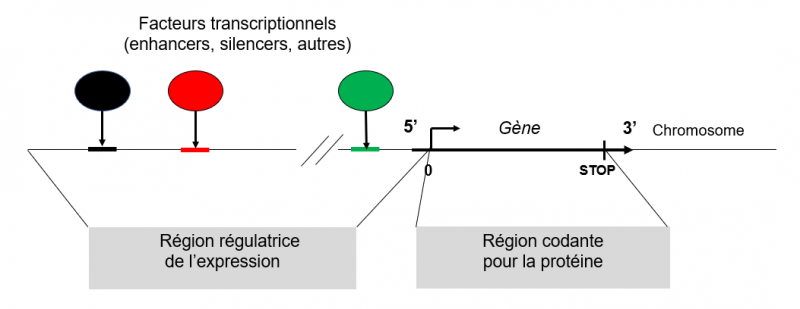
Un gène et sa région régulatrice : schéma très simplifié
Un gène est constitué d’une séquence qui code pour une protéine. A l’instar d’un véhicule, le gène peut être à l’arrêt ou en fonction. Lorsqu’il est en fonction (en cours d’expression), il produit la protéine pour laquelle il code de façon plus ou moins importante selon l’état de la cellule. Cette modulation de l’expression du gène est très étroitement contrôlée et met en jeu des facteurs transcriptionnels qui sont des protéines. En vert sur le schéma, le promoteur du gène est localisé en proximité immédiate du site d’initiation de la transcription (noté 0 sur le schéma, point à partir duquel est synthétisé l'ARN lors de la transcription). Son rôle est de contrôler l'expression de base du gène par le biais de facteurs transcriptionnels spécifiques. Mais d'autres sites de régulation peuvent exister à distance du promoteur en amont du site d’initiation de la transcription dans la "région régulatrice de l’expression" du gène. Certaines des protéines qui se fixent sur ces sites (noir et rouge sur le schéma) peuvent être des "enhancers" (activateurs) ou des "silencers" (ralentisseurs). Dans le cas de la lactase, c’est dans cette région régulatrice à distance du site d'initiation de la transcription que se trouve la mutation -13910:C>T (ou des autres, voir plus bas). Cette mutation permet l'activation du gène et entraîne la persistance de son expression chez l’adulte. Cette figure est en support de la suivante.
Pour le gène LCT, cette région se trouve très en amont du site d'initiation de la transcription, et plus précisément dans l'exon 13 du gène MCM6, dont il est adjacent sur le chromosome 2, comme il apparaît dans la figure ci-dessous.
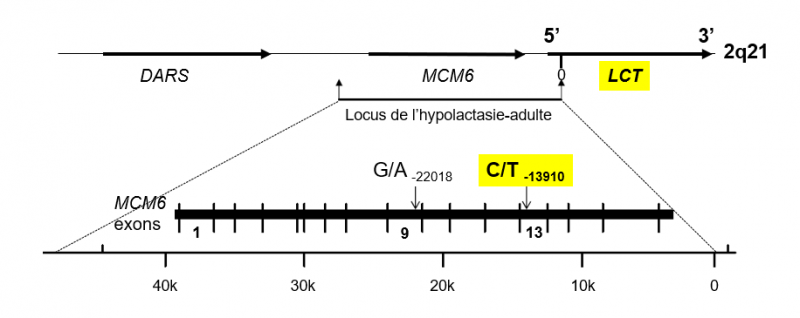
Les gènes DARS, MCM6 et Lactase (LCT)
Ces gènes sont localisés sur le chromosome 2 (locus 2q21). La séquence d'ADN du chromosome se prolonge de part et d'autre de la ligne du haut (ce chromosome comporte plus de 1200 gènes). Le site d'initiation de transcription du gène LCT est noté 0 (ligne du haut). On voit que la région régulatrice du gène LCT se prolonge dans le gène MCM6 dont les détails sont représentés en gras (ligne noire). Les séquences portant les mutations -13910:C>T et -22018:G>A sont signalées par les flèches dans les exons 13 et 9, respectivement. Adapté de Enattah et al.2002.
Pourquoi cette mutation -13910:C>T a-t-elle un impact aussi important sur l'activité du gène LCT ? L'activité transcriptionnelle de ce gène est complexe et la figure ci-dessous ne représente qu'une version simplifiée de la réalité.
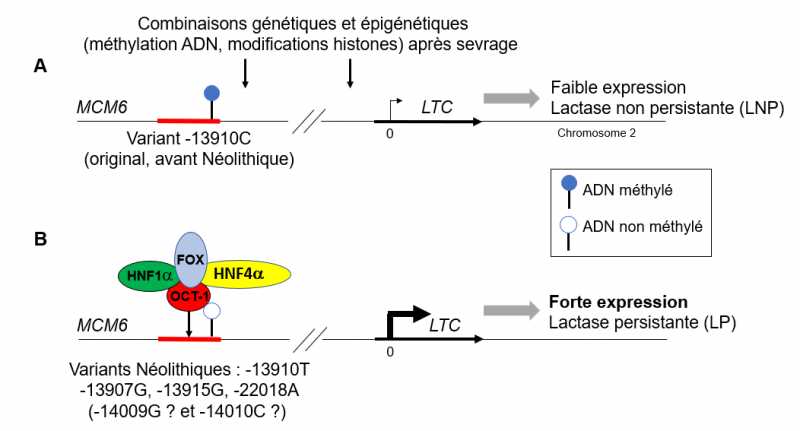
Mécanisme de modulation de l'expression du gène de la lactase (LCT). Schéma très simplifié
En A, la situation ancestrale dans laquelle des méthylations sur l'ADN (voir légende en encadré) empêchent l'activation de la transcription du gène LCT. Dans ces conditions, le complexe OCT-1/HNF1a/HNF4a/FOX ne peut exercer son effet activateur. L'expression du gène est très faible et conduit au trait de lactase non persistante (la norme chez les humains). En B, les séquences contenant la mutation -13910:T (trait rouge) échappent à l'inactivation épigénétique (du fait de la mutation ?) facilitant ainsi la persistance de l'expression du gène d'une part en évitant l'effet inhibiteur des méthylations et d'autre part en permettant l'effet "enhancer" (activateur) de la protéine OCT-1. L'expression du gène est forte et conduit au trait de lactase persistante. Adapté de Swallow et al. 2016; Labrie et al.2016; Kuchay 2020; Anguita-Ruiz et al. 2020.
Comme tous les gènes de notre génome, LCT est non seulement contrôlé par des facteurs transcriptionnels, mais également par des modifications épigénétiques, comme notamment (mais pas seulement) des méthylations de l'ADN sur certains nucléotides A et C. Sans entrer dans le détail, la méthylation de l'ADN peut fortement changer l'activité transcriptionnelle d'un gène sans en changer la séquence. Des études récentes (Labrie et al. 2016) ont fourni des preuves fonctionnelles de l'existence de ce mode complexe de régulation pour le gène de la lactase.
Dans la figure ci-dessus, la partie A présente la situation du gène dans la forme ancestrale (-13910:C) qui conduit au trait de lactase non persistante. Dans la partie B est représentée la situation lorsque la mutation -13910:T est présente. L'expression du gène n'est alors plus soumise aux effets inhibiteurs de la méthylation de l'ADN et la présence de la mutation permet la fixation du facteur transcriptionnel OCT-1 qui, au sein d'un complexe avec d'autres protéines HNF1a, HNF4a, et FOX, active la transcription du gène par son effet "enhancer" qui conduit au trait de lactase persistante.
Impact de ces mutations sur l'évolution vers le trait de lactase persistante
On a tout d'abord pensé que ce trait de lactase persistante était lié à une adaptation à la consommation de lait que les premiers agriculteurs et éleveurs ont introduit dans certaines populations humaines au Néolithique comme le stipule l'hypothèse "culture-historique" (voir plus loin).
En effet, la preuve la plus ancienne de l'existence d'une activité laitière remonte au septième millénaire avant J-C. Elle est basée sur des résidus organiques laitiers retrouvés dans les poteries au nord-ouest de l'Anatolie (emplacement actuel de la Turquie). Cette preuve d'activité laitière est associée à une forte concentration d'ossements de bovins, ovins et caprins dans les lieux de vie des humains de l'époque. L'analyse de ces ossements suggère que la gestion des produits laitiers a été concomitante à la domestication de ces animaux en Asie du Sud-Ouest au neuvième millénaire avant notre ère.
La pratique de l'élevage et de la transformation des produits laitiers a été apportée en Europe par les premiers agriculteurs via la côte méditerranéenne et en Europe Centrale via les Balkans. En Europe du Nord, des études approfondies sur les résidus organiques dans les tessons de poteries et le tartre dentaire montrent sans équivoque qu'au quatrième millénaire avant J-C, les habitants néolithiques des îles britanniques et de l'Irlande étaient des éleveurs laitiers accomplis. Il existe au Danemark et en Finlande préhistoriques des preuves de l'utilisation du lait parallèlement aux ressources aquatiques à partir de 4000-3000 avant J-C.
La fréquence de ce trait de lactase persistante varie considérablement d'une population à l'autre, allant de 5 % à près de 100 %, les fréquences les plus élevées étant observées chez les populations nord-européennes et certaines populations d'Afrique occidentale, d'Afrique orientale et du Moyen-Orient.
Une telle trajectoire de la fréquence des mutations conduisant du trait de lactase non persistante (la norme) vers le trait de lactase persistante (les mutants) témoigne d'une forte sélection naturelle en faveur de ce trait qui aurait débuté au Néolithique ancien.
Quel a été le moteur de cette sélection naturelle?
Deux hypothèses principales ont été considérées :
- L'hypothèse "culture-historique" selon laquelle le trait de lactase persistante était rare chez nos lointains ancêtres jusqu'à l'avènement de la production laitière au début du Néolithique. Sa fréquence a ensuite augmenté rapidement sous l'effet de la sélection naturelle, du fait de l'avantage nutritionnel apporté par la consommation de lait et des produits laitiers.
- L'hypothèse de la " cause inverse " selon laquelle la production laitière a été adoptée par des populations qui présentaient déjà des fréquences élevées du trait de lactase persistante. Pouvant naturellement consommer du lait sans dommage elles ont profité de cet avantage nutritionnel.
Cette deuxième hypothèse a toutefois été abandonnée lorsque les analyses de l'ADN ancien ont révélé l'absence de mutations conduisant au trait de lactase persistante chez les humains avant le Néolithique (Berger et al. PNAS2007).
Cependant, une grande incertitude demeure quant à aux facteurs qui ont contribué à cette évolution.
Des observations contre-intuitives
Si l'hypothèse "culture-historique" est la bonne, les populations qui dépendent fortement du lait (notamment les éleveurs nomades) devraient présenter des fréquences de lactase persistante plus élevées (à condition qu'elles ne soient pas passées récemment au pastoralisme et qu'elles ne se soient pas mélangées de manière importante avec des populations non pastorales). Mais ce n'est apparemment pas le cas.
En effet, chez les éleveurs de longue date d'Asie, comme les Mongols et les Kazakhs, la fréquence du trait lactase persistante observée est assez faible 12 % chez les Mongols et 24-30 % chez les Kazakhs.
Chez d'autres éleveurs d'Asie centrale, où le phénotype lactase persistante est bien corrélé avec la présence de la mutation -13.910:T, les fréquences lactase persistante se situaient entre 3% et 26%, avec une moyenne de 14% (Ségurel et al. 2017). De même, dans les populations tibétaines, les cinq mutations connues pour être associées au LP sont absentes, ce qui suggère soit une très faible fréquence du trait lactase persistante, soit une base génétique autre.
Par conséquent, en Asie, le tableau qui se dessine est celui d'une très faible fréquence de lactase persistante, que ce soit chez que les populations aient un mode de vie pastoral ou non.
Les bergers d'Asie centrale ne sont pas la seule exception au modèle attendu. En effet, les Sami, éleveurs de rennes en Scandinavie, ont une fréquence du trait lactase persistante plus faible que le reste de la population suédoise (40-75% contre 91%) malgré une plus grande dépendance au pastoralisme (60% contre 30%). De même, certaines ethnies pastorales africaines consommatrices de lait (50-90% de pastoralisme) ont des fréquences du trait de lactase persistante faibles comprises entre 3 et 25%.
Enfin, dans la région où les animaux ont été domestiqués pour la première fois (notamment en Turquie) et plus généralement autour de la Méditerranée, les populations qui utilisent le lait depuis des millénaires ont des fréquences du trait de lactase persistante modérées.
Cet ensemble de données n'est pas en faveur de l'hypothèse "culture-historique"
Des différences dans les facteurs culturels, écologiques, nutritionnels et environnementaux contribuent probablement aux quantités et aux quantités relatives de lait frais et transformé consommées dans les différentes populations humaines pourraient expliquer ces observations contre-intuitives.
Une autre explication ?
Une étude récente (Evershed et al. Nature 2022) publiée le 27 juillet 2022 dans la revue scientifique internationale Nature, a porté sur les mécanismes selon lesquels le trait de lactase persistante se serait mis en place durant le Néolithique. Cette étude a été réalisée par un consortium de plus de 100 chercheurs de diverses institutions académiques des pays suivants : Royaume Uni, Allemagne, France, Espagne, Pologne, Hongrie, République Tchèque, Pays Bas, Roumanie, Turquie, Ukraine, Autriche, Portugal, Italie, Finlande, et Etats Unis.
Dans cette étude, les chercheurs développent la thèse selon laquelle le trait de lactase persistante aurait été sélectionné sous la pression d'épisodes occasionnels et récurrents de famine et de maladies infectieuses qui auraient affecté de façon beaucoup plus drastique les porteurs du trait lactase non persistante, en termes de survie et de capacité de reproduction.
Ces chercheurs ont utilisé le matériel et les données recueillis sur plus de 550 sites archéologiques, pour étudier d'une part la présence de résidus de matière grasse d'origine laitière dans 13 000 tessons de poterie (marqueurs de la consommation de lait), et d'autre part la séquence de l'ADN d'anciens humains qui y vivaient (absence ou présence de mutations vers le trait de lactase persistante). Ces études s'appuyant sur les génomes de plus de 1700 anciens eurasiens ont montré que les premiers éleveurs étaient intolérants au lactose (absence des mutations génétiques conduisant au trait de lactase persistante) et que la tolérance au lait n'est devenue courante en Europe qu'après l'âge du bronze, il y a 5 000 à 4 000 ans avec l'apparition des mutations décrites dans les paragraphes précédents.
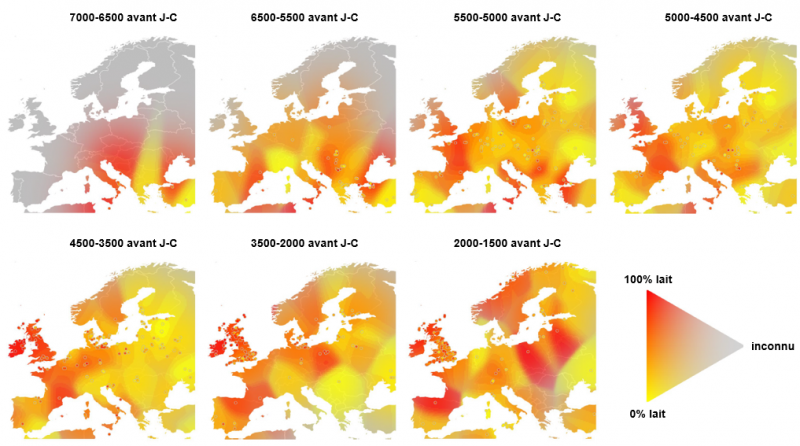
Variation régionale de l'utilisation du lait dans l'Europe préhistorique
Interpolation de la fréquence des résidus de matière grasse laitière dans les tessons de poterie (teinte) et de la confiance dans l'estimation (saturation de la couleur) en utilisant une estimation de densité bidimensionnelle. Les cercles en jaune ou en rouge (difficilement visibles sur la figure) indiquent les fréquences observées sur les sites de prélèvement. La large cline (variation phénotypique régulière et continue) sud-est à nord-est de saturation des couleurs au début de la période néolithique illustre un biais d'échantillonnage vers les preuves les plus anciennes de l'utilisation du lait. Une hétérogénéité substantielle dans l'exploitation du lait est évidente à travers l'Europe continentale. En revanche, dans les Îles britanniques et l'ouest de la France on observe un déclin progressif pendant 7000 ans après les premières preuves d'utilisation du lait vers 5500 avant J-C.
Adapté de Evershed et al. 2022.
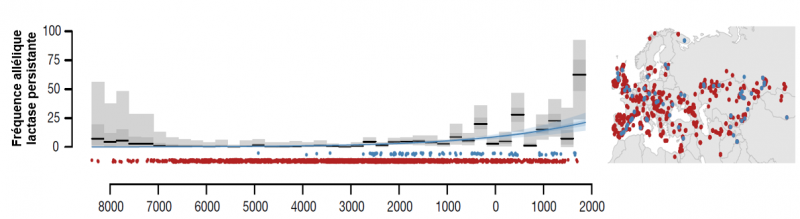
Variation régionale des fréquences des allèles lactase persistante dans l'Europe préhistorique
Données obtenues à partir du séquençage de l'ADN ancien (prélevé sur les restes humains de l'époque) sur le nucléotide en position -13910, c'est à dire -13910:C (trait de lactase non persistante, LNP) et -13910:T (trait de lactase persistante, LP). Les points bleus et rouges sur la carte et au dessus de l'échelle de temps, montrent respectivement la date et la localisation de 3 057 allèles LP et LNP provenant de 1 770 individus anciens. Les lignes noires à l'intérieur des barres grises représentent une estimation statistique de la fréquence des allèles LP dans chaque tranche de temps. Les barres grises montrent l'IC (intervalle de confiance) à 95 et 50% dans chaque tranche de temps. La ligne bleue est la courbe sigmoïde de maximum de vraisemblance du taux de fréquence des allèles LP en fonction du temps. L'ombrage bleu représente l'IC à 95 et 50 % estimé à partir des paramètres conjoints de la sigmoïde. L'échelle de temps de 8000 ans avant J-C et 2000 de notre ère est représentée au bas de la figure. Adapté de Evershed et al. 2022.
On voit sur la figure ci-dessus que le génotype lactase persistante (points et ligne bleus) n'est apparu que vers les années 5000 avant J-C pour s'amplifier jusqu'au moyen âge.
Pour tenter de comprendre l'origine de la capacité des européens à digérer le lait, les chercheurs ont modélisé la manière dont divers facteurs tels que la consommation de lait, les variations de taille et de densité des populations, la survenue de famines, les fluctuations dans l'exploitation des animaux sauvages ou domestiques, la survenue d'infections, pourraient moduler le trait de lactase persistante. Et les résultats - surprenants - ont montré qu'il y avait peu de corrélation entre le fait d'être tolérant au lactose et l'augmentation de la consommation de lait. Voir aussi le précédent paragraphe : "Des observations contre-intuitives".
L'image générale qui résulte de cette étude est que, si la production laitière a persisté tout au long de la période du Néolithique, son intensité a considérablement fluctué dans l'espace et dans le temps, ce qui laisse supposer des instabilités régionales spécifiques dans la production alimentaire et des changements culturels dans les préférences alimentaires. Ceci est cohérent avec des études antérieures qui ont montré des fluctuations régionales d'expansion et de ralentissement de la densité de population en Europe au cours de la même période (Shennan et al. Nat Comm 2013).
Les chercheurs ont alors tenté de savoir s'il existe encore aujourd'hui un lien entre la tolérance au lactose et la consommation de lait chez les Européens modernes. Dans la UK Biobank (Banque d'ADN constituée au Royaume Uni) qui regroupe des données sur la santé et la génomique d'un demi-million de personnes, ils ont constaté une faible corrélation entre la consommation de lait et la tolérance au lactose. Par exemple, 92 % des participants intolérants au lactose préfèrent le lait frais aux substituts. Autrement dit :
La tolérance au lactose ne présente aujourd'hui aucun avantage évident en matière de santé ou de fertilité, ce qui aurait pu sous-tendre la sélection naturelle dans le cadre de l'hypothèse "culture-historique".
Cela suggère donc que pour la grande majorité des personnes intolérantes au lactose, le coût de la consommation de lait n'est pas si élevé aujourd'hui, et ne l'était peut-être pas non plus au Néolithique. En effet, lorsqu'on est en bonne santé, un court épisode de diarrhée, de crampes abdominales, ou de flatulences est certes désagréable mais n'engage pas le pronostic vital. Compte tenu de l'exploitation préhistorique généralisée du lait et de ses effets relativement bénins chez des individus porteurs du trait de lactase non persistante en bonne santé aujourd'hui, les chercheurs proposent deux mécanismes connexes pour l'évolution du trait de lactase persistante:
- Selon le premier mécanisme dit "mécanisme de crise", les conséquences néfastes sur la santé de la consommation d'aliments à forte teneur en lactose par les personnes intolérantes se manifesteraient principalement pendant les famines, entraînant une sélection élevée mais épisodique en faveur du trait de lactase persistante. Ceci est dû au fait que la diarrhée induite par le lactose peut passer d'un état gênant à un état fatal chez les personnes gravement malnutries et que les produits laitiers à haute teneur en lactose (non fermentés) sont plus susceptibles d'être utilisés lorsque les autres ressources alimentaires sont épuisées. Ce "mécanisme de crise" prédit que les pressions de sélection conduisant au trait de lactase persistante auraient été plus fortes en période de famine.
- Selon le deuxième mécanisme dit "mécanisme chronique", l'augmentation de la charge pathogène - en particulier les zoonoses du fait de la promiscuité avec les animaux d'élevage - associée à l'agriculture et à l'augmentation de la densité et de la mobilité de la population aurait joué un rôle déterminant. La mortalité et la morbidité dues à l'exposition aux pathogènes auraient été amplifiées chez les porteurs du trait lactase non persistante chez qui les diarrhées et autres perturbations intestinales auraient représenté un lourd désavantage par rapport au porteur du trait lactase persistante. Ce "mécanisme chronique" prédit que les pressions de sélection pour le trait de lactase persistante auraient augmenté avec une plus grande exposition aux agents pathogènes.
Conclusion
Evershed et al. proposent que la sélection naturelle vers le trait de lactase persistante se soit accélérée pendant les périodes de stress nutritionnel sévère et d'infections, lorsque les personnes intolérantes au lactose étaient davantage susceptibles de mourir par rapport aux personnes ayant hérité des variants génétiques bénéfiques comme le -13910:T en Europe.
Critiques
Même si les résultats présentés par Evershed et al. sont généralement convaincants et leur analyse rigoureuse, leurs conclusions restent soumises à diverses critiques.
- Quelle a été l'ampleur et la portée des famines ou des épisodes de maladies infectieuses qui ont pu influencer l'évolution de la tolérance au lactose.
- Il faut également garder à l'esprit que les fromages et autres produits laitiers à faible teneur en lactose datent d'environ 7 400 ans en Europe et n'auraient pas eu représenté une aussi forte menace sur les porteurs du trait lactase non persistante.
- Pas sûr que cette étude explique entièrement pourquoi le trait de lactase persistante est absent, ou très rare, chez les premiers agriculteurs et certaines populations laitières de l'âge du bronze, qui ont pourtant dû être exposés également à la famine et aux infections.
- Comment la consommation de lait et de produits laitiers peut s'être répandue dans des endroits où la tolérance au lactose n'a jamais été habituelle, par exemple chez les Mongols.
- Enfin, le trait de lactase persistante s'est fortement développé en Europe et largement moins en Asie de l'Est où les famines et les infections ont dû être également présentes pendant toute la période du Néolithique.
Bibliographie
- Anguita-Ruiz et al. Genetics of Lactose Intolerance: An Updated Review and Online Interactive World Maps of Phenotype and Genotype Frequencies. Nutrients 2020
- Burger et al. Absence of the lactase-persistence-associated allele in early Neolithic Europeans. Proc Natl Acad Sci 2007
- Enattah et al. Identification of a variant associated with adult-type Hypolactasia. Nature Genetics 2002
- Evershed et al. Dairying, diseases and the evolution of lactase persistence in Europe. Nature 2022
- Fong et al. The human lactase persistence-associated SNP −13910*T enables in vivo functional persistence of lactase promoter reporter transgene expression. Human Genetics 2012
- Gerbault et al. How Long Have Adult Humans Been Consuming Milk? International Union of Biochemistry and Molecular Biology 2013
- Itan et al. A worldwide correlation of lactase persistence phenotype and genotypes. BMC Evolutionary Biology 2010
- Kuchay. New insights into the molecular basis of lactase non-persistence/ persistence: a brief review. Drug Discoveries & Therapeutics 2020
- Labrie et al. Lactase non-persistence is directed by DNA variation-dependent epigenetic aging. Nat Struct Mol Biol. 2016
- Leseva et al. Differences in DNA Methylation and Functional Expression in Lactase Persistent and Non-persistent Individuals. Scientific Reports 2018
- Lewinsky et al. T-13910 DNA variant associated with lactase persistence interacts with Oct-1 and stimulates lactase promoter activity in vitro. Human Molecular Genetics 2005
- Olds et al. –13915*G DNA polymorphism associated with lactase persistence in Africa interacts with Oct-1. Human Genetics 2011
- Porzi et al. Development of Personalized Nutrition: Applications in Lactose Intolerance Diagnosis and Management. Nutrients 2021
- Ségurel and Bon. On the Evolution of Lactase Persistence in Humans. Annual Review of Genomics and Human Genetics 2017
- Shennan et al. Regional population collapse followed initial agriculture booms in mid-Holocene Europe. Nature Communications 2013
- Swallow and Troelsen. Escape from epigenetic silencing of lactase expression is triggered by a single-nucleotide change. Nature Structural & Molecular Biology 2016
- Troelsen. Adult-type hypolactasia and regulation of lactase expression. Biochimica et Biophysica Acta 2005
Date de dernière mise à jour : 30/08/2022
Ajouter un commentaire