
Entrée du virus dans les cellules
Récepteur du SARS-CoV-2 et conséquences thérapeutiques
Résumé
La mise au point de thérapies spécifiques et d’un vaccin contre le SARS-CoV-2, le coronavirus de Wuhan responsable de l’épidémie Covid-19, représente une priorité. Dans cette optique, connaître le mécanisme par lequel le virus entre dans les cellules est fondamental. Les données déjà obtenues sur le SARS-CoV-1 impliqué dans l’épidémie de SARS qui toucha la chine en 2002-2003 et celles obtenues plus récemment sur le SARS-CoV-2, montrent que l’entrée du virus dans les cellules se fait en deux étapes :
Etape 1: Liaison de la protéine virale "spike" (protéine S) à une protéine des membranes cellulaires appelée ACE2 (enzyme de conversion de l’angiotensine).
Etape 2: Activation de la protéine S par des protéases cellulaires dont notamment la protéine TMPRSS2. Cette protéase joue un rôle majeur en activant l’entrée du virus dans les cellules.
Conséquences:
- L'inhibition de TMRPSS2 pourrait représenter une stratégie thérapeutique pour bloquer l’infection Covid-19.
- Des études récentes ont montré qu’un médicament, le Camostat Mesylate, utilisé au Japon pour le traitement des pancréatites chroniques, bloque la protéase TMPRSS2 et pourrait être utilisé pour traiter les malades atteints de Covid-19.
- Un essai clinique va être lancé au Japon pour tester cette hypothèse en Avril 2020. Le Nafamostat Mesylate analogue du Camostat sera également testé.
- La chloroquine exerce son effet antiviral en bloquant l'entrée du virus par la voie des endosomes.

Explications
Nos connaissances actuelles sur le processus d’entrée du SARS-CoV-2 sont largement issues de ce que l’on sait sur le SARS-CoV-1 responsable de l’épidémie de SRAS qui toucha la Chine en 2002-2003 et « cousin » du SARS-CoV-2 (Masters 2006 ; Simmons et al. 2013). Les études toutes récentes menées sur le SARS-CoV-2 (Hoffmann et al. 2020 ; Liu et al. 2020 ; Letko et al. 2020 ; Zhang et al. 2020 ; Yan et al. 2020) confirment et viennent en support de celles réalisées précédemment sur le SARS-CoV-1.
Brève description du virus SARS-CoV-2
La particule virale a une forme quasi sphérique et un diamètre compris entre 80 et 120 nm (rappel : 1 nm représente un millième de micron qui représente lui-même un millième de mm). Elle est constituée d’une enveloppe elle-même constituée d’une bicouche lipidique (issue de la cellule qui a produit le virus) et de 3 protéines, les protéines S (skipe), M (membrane) et E (enveloppe). Au sein de la particule virale se trouve le génome viral, ARN simple brin positif (c’est à dire codant) étroitement lié à une protéine la nucléocapside (N). L’assemblage nucléocapside-ARN se fait un peu à la façon dont des perles (N) sont attachées sur un fil (l’ARN).
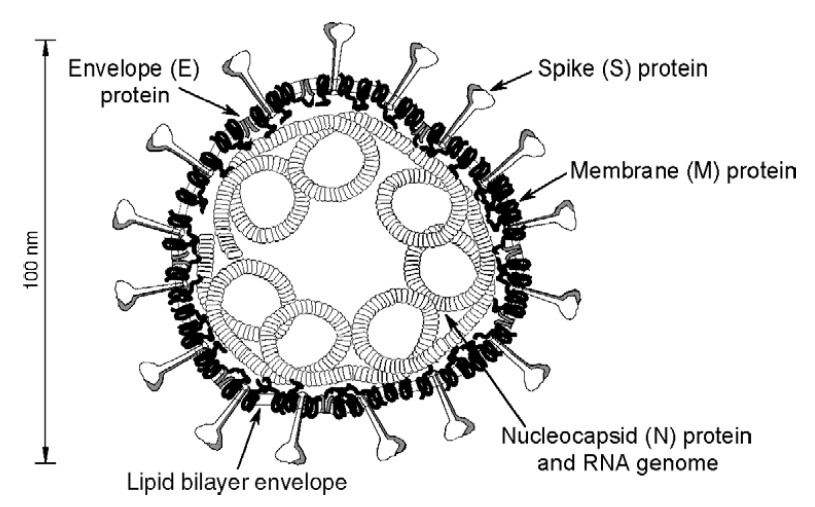
Schéma du virus SARS-CoV (Masters 2006)
Le génome du SARS-CoV-2 (entre 27 et 32 kb) code pour au moins 27 protéines, incluant 15 protéines non-structurales, 4 protéines structurales et 8 protéines dites auxiliaires.
Le virus ne peut se multiplier spontanément à l’extérieur des cellules car il ne possède pas la machinerie nécessaire (ribosomes, ARNs de transfert et nombreux autres agents) pour la synthèse des protéines. Celle-ci lui est fournie par les cellules. L’entrée du virus dans les cellules est donc une première étape essentielle à sa survie.
Entrée du virus SARS-CoV-2 dans la cellule
L’entrée du virus dans la cellule est un processus complexe impliquant deux étapes principales.
Première étape: fixation du virus sur le récepteur
Au cours de la première étape, le domaine de liaison au récepteur (RBD, receptor binding domain) de la protéine S (skipe) se fixe avec une très forte affinité sur le domaine peptidase (PD, peptidase domain) de l’enzyme de conversion de l’angiotensine 2 (ACE2, angiotensin converting enzyme 2), une des protéines de la membrane cellulaire. La protéine S est schématisée dans la figure ci-dessous.
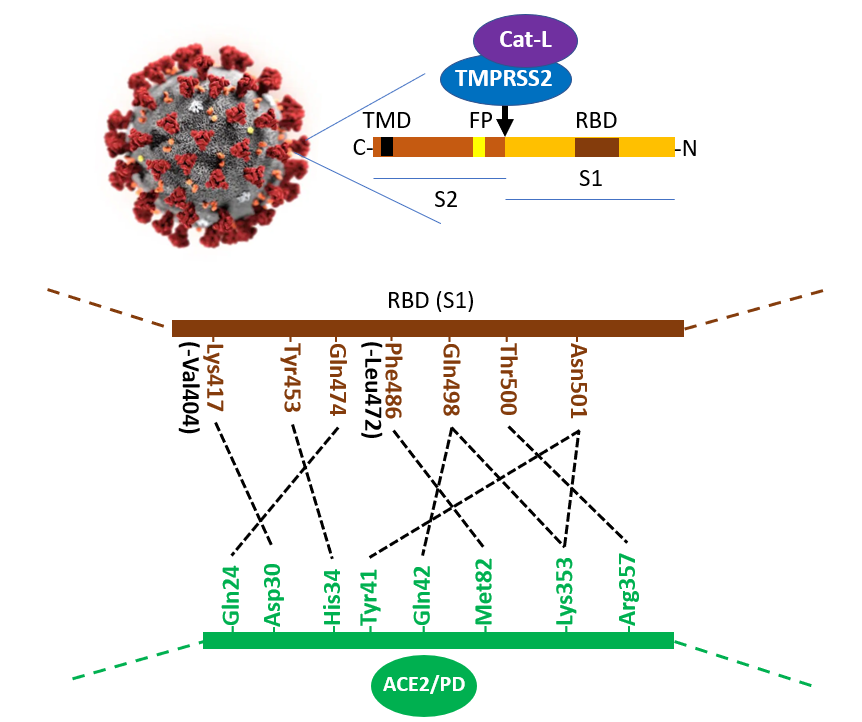
La protéine skype (S) et interaction entre le domaine de liaison du récepteur (RBD) de S et le domaine peptidase (PD) de ACE2.
Partie haute: le virus et vue schématique de la protéine S, montrant le domaine transmembranaire (TMD), le peptide de fusion (FP) et le domaine de liaison du récepteur (RBD). La flèche noire représente le site de coupure de S par les protéases TMPRSS2 et Cathepsine-L (Cat-L).
Partie basse: vue schématique de l'interaction entre le domaine de liaison du récepteur (RBD) de S de SARS-CoV-2 et le domaine peptidase (PD) de ACE2. Les pointillés vert et marron figurent les parties restantes des deux protéines. Les pointillés noirs figurent les interactions entre le RBD et ACE2. Les acides aminés en noir, sont ceux correspondants à la protéine S du SARS-CoV-1 et différents de ceux présents dans la protéine S du SARS-CoV-2. (Yan et al. 2020).
La protéine S est schématiquement divisée en deux parties S1 et S2. La partie S1 de la protéine comprend le domaine de liaison au récepteur (RBD). La partie S2 est impliquée dans l’entrée proprement dite dans la cellule, par un processus de fusion membranaire qui dépend du peptide de fusion (FP, fusion peptide) ; cette partie comporte également le domaine transmembranaire (TMD, transmembrane domain) qui ancre la protéine S dans la membrane virale.
La protéine ACE2 est présente dans le poumon (pneumocytes et macrophages), le cœur, l’intestin et le rein. La forte affinité de fixation du virus sur ACE2 résulte d’interactions polaires et de ponts hydrogène entre des acides aminés de la protéine virale S et des acides aminés de la protéine cellulaire ACE2. La figure ci-dessus (partie basse) montre les acides aminés impliqués dans cette fixation (Yan et al. 2020).
Lorsqu’on compare la séquence du RBD du virus SARS-CoV-1 avec celle du SARS-CoV-2 des différences apparaissent. Rappelons que l’homologie de séquence pleine longueur en acides aminés entre les deux coronavirus est de 94,6%. Notamment dans le SARS-CoV-1 les acides aminés Val404 et Leu472 remplacent les Lys417 et Phe498 du SARS-CoV-2 (Yan et al. 2020). On comprend ainsi pourquoi l’affinité de liaison entre les deux virus et la protéine ACE2 sera différente. Or, l’affinité entre la protéine virale et la protéine récepteur humaine (ACE2) joue un rôle déterminant dans la transmissibilité du virus. Si pour un coronavirus d’une espèce animale X cette affinité avec ACE2 est faible (du fait de différences dans la séquence du RBD), il y a peu de chance que l’espèce X soit responsable de la transmission à l’homme. C’est le cas du coronavirus MERS (transmis par le chameau) qui utilise en fait un autre récepteur, différent de ACE2. C’est sur cette base que se pose la question de savoir quelle est l’espèce animale relai entre la chauve-souris et l’homme dans l’infection. C’est également sur cette base que le pangolin avait été suspecté être cette espèce animale. En effet, l’homologie de séquence du domaine de liaison (RBD) au récepteur du coronavirus du pangolin est 99% identique à celle du SARS-CoV-2 (Xiao et al. 2020). Cependant, d’autres études ( Liu et al. 2020, Zhang et al. 2020, Lam et al. 2020) ont ensuite montré que si on considère la séquence pleine longueur (et non seulement la séquence du RBD) du coronavirus qui infecte les pangolins de Malaisie, l’homologie de séquence n’est plus que de l’ordre de 85 à 92% et cela suggère donc que la source du SARS-CoV-2 ne peut être cet animal.
Deuxième étape: activation de l'entrée cellulaire
Bien que l’ACE2 soit présente dans les cellules endothéliales de tous les organes, le SARS-CoV est essentiellement pathogène pour le poumon. De plus, le tropisme cellulaire du virus ne corrèle pas strictement avec la distribution de ACE2. Par exemple, alors que l’expression de ACE2 se fait dans les pneumocytes 1 et 2, seuls les pneumocytes 1 impliqués dans les échanges d’oxygène sang-tissu au niveau des alvéoles pulmonaires sont infectés par le virus. Cela suggère que le récepteur ne peut agir seul et qu’un cofacteur est nécessaire pour l’entrée du virus dans les pneumocytes (Kawase et al. 2012).
Pour le SARS-CoV, ce rôle de cofacteur est tenu par au moins deux protéases, la TMPRSS2 (transmembrane serine protéase) et la cathepsine-L (une cystéine protéase).
Lorsque la protéine S est fixée sur l’ACE2, un changement de conformation de la protéine se produit, rend celle-ci sensible aux protéases TMPRSS2 et/ou cathepsine L qui par coupure éliminent la partie S1. Il s’agit d’un processus d’activation protéolytique de la protéine virale S qui « découvre » le peptide de fusion FP. C’est ce peptide FP qui, rendu accessible, permet l’entrée proprement dite du virus dans la cellule soit par un processus de fusion entre la membrane virale et la membrane cellulaire après activation par la TMPRSS2, soit par un processus de fusion entre la membrane virale et la membrane des endosomes cellulaires après activation par la cathepsine-L (Simmons 2013). Ce processus se termine par l’introduction du génome viral dans la cellule comme indiqué dans la figure ci-dessous. Il semble que ces deux processus ne soient pas mutuellement exclusifs et agissent simultanément.
A noter que la chloroquine reconnue pour son effet antiviral sur le SARS-CoV-2 (Wang et al. 2020 et Gao et al. 2020) exercerait son action en augmentant le pH dans les endosomes, réduisant ainsi l'activité de la cathepsine-L et inhibant la fusion de la membrane virale et de la membrane des endosomes.
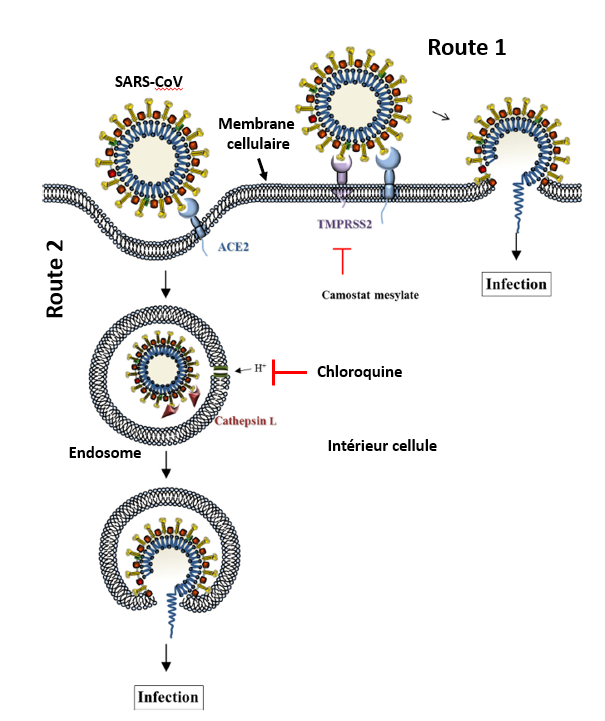
Schéma du mécanisme d'entrée du virus SARS-CoV dans les cellules (selon Simmons 2013)
La libération du génome viral dans la cellule peut se faire suivant deux routes différentes, suivant la protéase utilisée.
Route 1 : activation de la protéine virale S par la protéase TMPRSS2. La protéine TMPRSS2 est exprimée sur les membranes comme la protéine ACE2. Ces deux protéines cohabitent et la liaison de la protéine S du virus sur ACE2 suffit pour que TMRPSS2 coupe S, exposant ainsi le peptide de fusion FP. C’est par l’intermédiaire de FP que la membrane du virus fusionne avec celle de la cellule. La particule virale ainsi ouverte peut libérer son contenu, c’est à dire le génome viral, dans la cellule.
Route 2 : activation de la protéine virale S par la cathepsine-L. Les endosomes se forment par invagination de la membrane cellulaire, suite à la liaison du virus sur son récepteur. Dans les endosomes, le pH diminue par entrée de protons. Ce processus conduit à l’activation de la cathepsine-L qui peut alors couper et activer la protéine S du virus, exposant ainsi le peptide de fusion FP. C’est par l’intermédiaire de FP que la membrane du virus fusionne avec celle de l’endosome. La particule virale ainsi ouverte peut libérer son contenu, c’est à dire le génome viral, dans la cellule.
Il est possible que d’autres protéases soient également impliquées dans l’activation de la protéine virale S (Simmons 2013).
Conséquences thérapeutiques
Les études décrites ci-dessus montrent clairement comment le clivage protéolytique de la protéine virale S est essentiel pour l’entrée du virus dans les cellules. La connaissance de ce mécanisme ouvre la voie à la mise en place de stratégies thérapeutiques. Ces stratégies visent les acteurs essentiels de l’entrée virale, c’est à dire les protéases TMPRSS2 et cathepsine-L. Du fait du rôle central que joue la cathepsine-L dans le fonctionnement des endosomes, son inhibition risque de provoquer des effets délétères et une application clinique semble pour le moment peu probable. Mais l'inhibition indirecte de cette protéase par la chloroquine qui augmente le pH à l'intérieur des endosomes, semble avoir un effet bénéfique sur l'infection Covid-19 (Service D Raoult, CHU La Timone). Pour ce qui concerne la protéase TMPRSS2, il existe déjà un médicament utilisé au Japon pour le traitement de la pancréatite chronique, le camostat mesylate, qui bloque l’activité biologique de la TMPRSS2 (Kawase et al. 2012 ; Zhou et al. 2015; Hoffmann et al. 2020). Ce médicament et certaines molécules apparentées (notamment le nafamostat mesylate) aux mêmes effets thérapeutiques pourraient être utilisés pour le traitement des malades infectés par le SARS-CoV-2 dans les prochaines semaines (https://www.nippon.com/en/news/yjj2020031800741/pancreatitis-drug-may-block-coronavirus-infection-u-of-tokyo.html).
De plus, on sait que les malades guéris de l’infection SARS-CoV-1 possèdent des anticorps neutralisants qui peuvent être détectés deux ans après l’infection et qui sont dirigés contre le protéine virale S. Il a été récemment montré que ces anticorps sont capables de bloquer l’infection in vitro de cellules humaines par le SARS-CoV-2 (Hoffmann et al. 2020). Un vaccin dirigé contre la protéine virale S du SARS-CoV-1 pourrait donc être efficace pour une protection contre le SARS-CoV-2.
Date de dernière mise à jour : 14/11/2020
Commentaires
-
I'm really enjoying the design and layout of your website.
It's a very easy on the eyes which makes it much more enjoyable for me
to come here and visit more often. Did you hire out a designer
to create your theme? Outstanding work! -
I'm not sure why but this blog is loading incredibly slow for
me. Is anyone else having this issue or is it a issue on my end?
I'll check back later on and see if the problem still exists. -
Amazing blog! Do you have any helpful hints for aspiring writers?
I'm hoping to start my own website soon but I'm a little lost on everything.
Would you suggest starting with a free platform like Wordpress or go
for a paid option? There are so many options out there
that I'm totally overwhelmed .. Any recommendations?
Thanks! -
Heya i am for the first time here. I came across this board and I find It really useful &
it helped me out much. I hope to give something back and help
others like you aided me. -
I'm not sure why but this website is loading incredibly slow
for me. Is anyone else having this problem or is it a issue on my end?
I'll check back later and see if the problem still exists. -
Way cool! Some very valid points! I appreciate you penning this write-up plus the rest of the website is extremely good.
-
First off I want to say great blog! I had a quick question that I'd like to ask if you don't mind.
I was interested to know how you center yourself and clear your thoughts before writing.
I have had a difficult time clearing my mind in getting my ideas out there.
I truly do take pleasure in writing however it just seems like the first 10 to 15 minutes
are generally lost simply just trying to figure out how to begin. Any
suggestions or hints? Many thanks! -
Hello there, You've done a great job. I'll definitely digg it and personally
suggest to my friends. I am sure they'll be
benefited from this site. -
Hi there just wanted to give you a quick heads up.
The words in your post seem to be running off the screen in Firefox.
I'm not sure if this is a formatting issue or something to do with web browser
compatibility but I figured I'd post to let you know.
The design look great though! Hope you get the problem fixed soon.
Many thanks -
This design is wicked! You definitely know how to keep a reader amused.
Between your wit and your videos, I was almost moved
to start my own blog (well, almost...HaHa!) Wonderful job.
I really enjoyed what you had to say, and more than that, how you presented it.
Too cool! -
Incredible story there. What occurred after? Thanks!
-
I do not know if it's just me or if perhaps everyone else experiencing
problems with your site. It looks like some of the text within your posts are running
off the screen. Can somebody else please provide feedback and
let me know if this is happening to them as well?
This could be a issue with my web browser because I've had this happen previously.
Thanks -
Its like you read my mind! You seem to know so much about this, like you wrote the
book in it or something. I think that you could do with some pics to
drive the message home a little bit, but instead of
that, this is fantastic blog. A fantastic read. I will certainly be back. -
It's in fact very complex in this full of activity life
to listen news on TV, thus I just use internet for that reason, and get the latest news. -
Great delivery. Solid arguments. Keep up the amazing work.
-
Heya i am for the first time here. I found this board and I find It really useful
& it helped me out a lot. I hope to give something back and help others like
you aided me. -
Having read this I believed it was rather informative.
I appreciate you taking the time and energy to put this information together.
I once again find myself spending a lot of time both reading and posting comments.
But so what, it was still worthwhile! -
Hello, I would like to subscribe for this website to take most recent updates,
thus where can i do it please assist. -
Sigurno vam se barem jednom dogodilo da potpiše ugovor s poslodavcem koji
obećava brda i doline, a na kraju ne isplati plaću.
Na današnjem pretrpanom i kaotičnom tržištu, nemoguće je znati
kome uistinu možete vjerovati. Zato su lažne reklame postale naša
svakodnevica, a jedini način da se to spriječi je kontinuirana edukacija i provjera.
U takvim trenucima ključni alat za svakog kupca postaju specijalizirane platforme za recenzije.
Prije nego što potrošite svoj novac ili predate životopis u
neku firmu, svakako posjetite iskustva recenzije.
Baza podataka sadrži iskrene ocjene o obrtima, tvrtkama i lokalnim majstorima, što vam drastično
olakšava pretragu.
Vaša osobna priča također može spasiti nekoga od loše investicije.
Nakon što završite suradnju s nekom firmom, odvojite minutu vremena
i napišete kratku recenziju. Time stvaramo pritisak na tržište da podigne kvalitetu usluga, i zajednički gradimo transparentnije poslovno okruženje za sve nas. -
Hey! Do you use Twitter? I'd like to follow you if that would be okay.
I'm definitely enjoying your blog and look forward
to new posts. -
Highly descriptive blog, I liked that bit. Will there be
a part 2? -
What's up, of course this article is genuinely pleasant and I have learned lot of things from it on the topic of blogging.
thanks. -
I'm really enjoying the design and layout of your site.
It's a very easy on the eyes which makes it much more enjoyable
for me to come here and visit more often. Did you hire out a designer to
create your theme? Excellent work! -
My family all the time say that I am killing my time
here at net, however I know I am getting knowledge all the time by reading thes nice articles
or reviews. -
This blog was... how do I say it? Relevant!!
Finally I have found something that helped me. Kudos! -
Hello! This is kind of off topic but I need some advice from an established blog.
Is it very hard to set up your own blog? I'm not very techincal but I can figure
things out pretty fast. I'm thinking about creating my own but I'm not sure where to start.
Do you have any ideas or suggestions? Appreciate it -
Thanks for sharing your info. I truly appreciate your efforts and I will be waiting for your
next write ups thank you once again. -
Great article, thanks for sharing.
Reference: https://www.pexels.com/vi-vn/@11bet-ing-2162387102/ -
Greetings! This is my first visit to your blog! We are a collection of volunteers and starting a new
project in a community in the same niche. Your blog provided
us valuable information to work on. You have done a outstanding job! -
I really like reading through an article that will make men and women think.
Also, thanks for allowing me to comment! -
Nice post. I was checking continuously this blog and I am impressed!
Extremely helpful information specifically the last part :) I care for such information much.
I was seeking this certain information for a very long time.
Thank you and best of luck. -
Good blog post. I definitely love this site. Thanks!
-
Appreciate the recommendation. Will try it out.
-
You actually make it seem so easy with your presentation but I find this matter to be really something that I think I would never understand.
It seems too complicated and very broad for
me. I am looking forward for your next post, I'll try to get the
hang of it! -
Hi there everyone, it's my first go to see at this web site, and paragraph is
genuinely fruitful in support of me, keep up posting these articles. -
I don't even know how I ended up here, but I believed this publish was great.
I do not recognise who you might be however certainly you
are going to a well-known blogger if you aren't already.
Cheers! -
Salam hangat untuk seluruh anggota forum. Senang bisa bergabung dan berdiskusi bersama.
-
It's actually a great and useful piece of information. I
am happy that you shared this helpful info with us.
Please stay us up to date like this. Thank you for
sharing. -
I visited multiple sites but the audio quality for audio songs
current at this web site is truly marvelous. -
Wonderful blog! Do you have any hints for aspiring writers?
I'm planning to start my own blog soon but I'm a little
lost on everything. Would you suggest starting with a free
platform like Wordpress or go for a paid option? There are so many choices out there that I'm completely confused ..
Any recommendations? Thanks! -
It's remarkable in support of me to have a web page, which is beneficial for my knowledge.
thanks admin -
Hello just wanted to give you a brief heads up
and let you know a few of the pictures aren't loading properly.
I'm not sure why but I think its a linking issue. I've tried it in two
different browsers and both show the same results. -
Good post. I definitely appreciate this site.
Keep it up! -
Whats up this is kinda of off topic but I was wanting to know if blogs use WYSIWYG editors or if you have to manually code with HTML.
I'm starting a blog soon but have no coding skills so I wanted to get guidance from someone with experience.
Any help would be enormously appreciated! -
Yesterday, while I was at work, my cousin stole my
apple ipad and tested to see if it can survive a 30 foot drop, just so she can be
a youtube sensation. My iPad is now broken and she has 83 views.
I know this is totally off topic but I had to share it with someone! -
Thanks for finally writing about >entrée du SARS-CoV-2 dans les cellules <Liked it!
-
We are a group of volunteers and starting a new scheme in our community.
Your website provided us with valuable info to work on. You've done a formidable job and our whole community will be grateful to you. -
Its not my first time to go to see this website, i am browsing this site dailly and get pleasant data from here every day.
-
Wow, this piece of writing is pleasant, my younger sister is analyzing these things, so I am going to inform her.
-
This blog was... how do you say it? Relevant!!
Finally I've found something that helped me. Cheers! -
It's not my first time to go to see this site, i am browsing this site
dailly and get fastidious data from here daily. -
When someone writes an piece of writing he/she keeps the plan of a user in his/her brain that how a user can be
aware of it. Therefore that's why this article
is outstdanding. Thanks! -
Good day! I could have sworn I've visited your blog before but after looking at some
of the posts I realized it's new to me. Anyways, I'm definitely
pleased I stumbled upon it and I'll be bookmarking it and checking back often! -
Heya i'm for the first time here. I came across this board and I find
It really useful & it helped me out much. I hope to give something back and help others like you helped me. -
At this moment I am going to do my breakfast, later than having my breakfast
coming again to read other news. -
Good article. I'm experiencing many of these issues as well..
-
Postingan yang sangat informatif! Informasi ini sangat relevan bagi
siapa pun yang ingin memahami perkembangan industri
migas di tanah air. Penting bagi kita untuk terus mendukung dan mengenal lebih dekat perusahaan legendaris seperti **PT.
ARUN NGL** yang telah menjadi bagian dari sejarah besar kemajuan energi tanah air.
Terima kasih sudah berbagi! Kunjungi PT. ARUN NGL -
Way cool! Some very valid points! I appreciate you
penning this write-up plus the rest of the site is really good. -
Heya i'm for the primary time here. I came across this board and I find It really helpful & it
helped me out much. I hope to provide something back and help others like you aided me. -
Pretty! This has been a really wonderful article.
Thank you for providing this info. -
Inspiring story there. What occurred after? Take care!
-
What's Happening i am new to this, I stumbled upon this I've
found It absolutely useful and it has aided me out loads.
I'm hoping to give a contribution & aid different customers like its
aided me. Great job. -
I’m not that much of a online reader to be honest but your sites really
nice, keep it up! I'll go ahead and bookmark your website to come back down the road.
All the best -
What's up, just wanted to say, I enjoyed this article. It was funny.
Keep on posting! -
Useful information. Lucky me I discovered your web site unintentionally,
and I'm stunned why this coincidence did not came about in advance!
I bookmarked it. -
Excellent, what a website it is! This webpage presents valuable facts to us, keep it
up. -
Hey there, I think your blog might be having browser compatibility issues.
When I look at your blog site in Firefox, it looks
fine but when opening in Internet Explorer, it has some overlapping.
I just wanted to give you a quick heads up! Other then that, great blog! -
I wanted to thank you for this excellent read!! I certainly enjoyed every bit of it.
I have you book marked to check out new things you post… -
Hmm it seems like your site ate my first comment (it was super long)
so I guess I'll just sum it up what I submitted and say, I'm thoroughly enjoying your blog.
I as well am an aspiring blog blogger but I'm still new to everything.
Do you have any tips and hints for rookie blog writers?
I'd definitely appreciate it. -
It's in fact very difficult in this busy life to listen news on Television, thus I just use world
wide web for that purpose, and take the most up-to-date news. -
Thanks for finally talking about >entrée du
SARS-CoV-2 dans les cellules <Liked it! -
Today, while I was at work, my sister stole my iPad and tested to see if it can survive
a 40 foot drop, just so she can be a youtube sensation. My iPad is now destroyed and she has
83 views. I know this is totally off topic but I had to share it with someone! -
Hi there! This blog post couldn't be written any better!
Going through this post reminds me of my previous roommate!
He continually kept talking about this. I will forward this article to him.
Pretty sure he will have a good read. Thank
you for sharing! -
Howdy this is kind of of off topic but I was wanting to know if blogs use WYSIWYG editors or if you have to manually code with HTML.
I'm starting a blog soon but have no coding know-how so I wanted to get advice
from someone with experience. Any help would be enormously appreciated! -
Hello, just wanted to tell you, I enjoyed this post. It was practical.
Keep on posting! -
We're a bunch of volunteers and opening a brand new scheme in our community.
Your web site offered us with valuable info to work
on. You've done an impressive activity and our entire community can be grateful to you. -
An impressive share! I've just forwarded this onto a colleague who was
doing a little homework on this. And he actually bought me dinner due to the fact that I
found it for him... lol. So let me reword this.... Thank YOU for the meal!!
But yeah, thanx for spending the time to discuss this
topic here on your web page. -
It's awesome to visit this web page and reading the views
of all colleagues on the topic of this post, while I am also
eager of getting familiarity. -
Greetings from California! I'm bored to tears at
work so I decided to browse your website on my iphone during lunch break.
I love the knowledge you present here and can't wait to take a look when I get home.
I'm amazed at how fast your blog loaded on my phone ..
I'm not even using WIFI, just 3G .. Anyhow, wonderful blog! -
I feel this is among the so much important information for me.
And i am satisfied studying your article. However wanna observation on few normal
issues, The web site taste is wonderful, the articles is truly nice :
D. Just right task, cheers -
It is not my first time to go to see this web site, i am browsing this web site dailly and get pleasant
data from here all the time. -
My brother recommended I would possibly like this blog.
He used to be entirely right. This put up actually made my day.
You cann't consider simply how a lot time I had spent for this info!
Thank you! -
Hey there! This is my first visit to your blog! We are a group of volunteers and
starting a new initiative in a community in the same niche.
Your blog provided us beneficial information to work on. You have done a wonderful job! -
Useful info. Lucky me I discovered your site accidentally, and I am stunned why this coincidence did not happened earlier!
I bookmarked it. -
Hello! Do you know if they make any plugins to protect
against hackers? I'm kinda paranoid about losing everything I've worked hard on. Any suggestions? -
Hi there, i read your blog occasionally and i own a similar one
and i was just curious if you get a lot of spam comments? If so how do you reduce
it, any plugin or anything you can suggest?
I get so much lately it's driving me crazy so any help is very much appreciated. -
You actually make it seem so easy with your presentation but I find this topic to be really something which I think I would never understand.
It seems too complex and extremely broad for me. I am looking forward for your
next post, I'll try to get the hang of it! -
I like the valuable info you provide to your articles.
I'll bookmark your weblog and test again right here regularly.
I am fairly certain I'll be told plenty of new stuff right right
here! Good luck for the following! -
You are so interesting! I do not believe I've read through something like that before.
So nice to discover somebody with some unique thoughts on this subject.
Really.. thanks for starting this up. This web site is something that is required on the internet, someone with a bit of originality! -
Hello, this weekend is nice designed for me, since this time i am reading this wonderful educational piece of writing here at my home.
-
Its like you read my mind! You appear to know a lot about this, like you wrote the book in it or something.
I think that you could do with some pics to drive the message home a little bit, but instead of that, this is
great blog. A fantastic read. I'll definitely be back. -
I got this website from my friend who told me about this website and now this time I am browsing this web page
and reading very informative articles or reviews here. -
Your mode of explaining everything in this piece of writing is in fact good, every one be able to without
difficulty understand it, Thanks a lot. -
Right away I am going away to do my breakfast, after having my breakfast coming over again to
read further news. -
Woah! I'm really enjoying the template/theme of
this site. It's simple, yet effective. A lot of times it's challenging to get
that "perfect balance" between superb usability and visual appearance.
I must say you have done a superb job with this.
Additionally, the blog loads very quick for me on Opera.
Exceptional Blog! -
It's going to be ending of mine day, however before end I am reading this fantastic paragraph to improve my knowledge.
-
Everything is very open with a clear description of the issues.
It was definitely informative. Your site is very helpful.
Many thanks for sharing! -
Un punto a considerar es la riesgo de cada tragamoneda.
Las tragamonedas tranquilas reparten premios chicos pero frecuentes, óptimas para
jugadas extensas. -
Howdy! I just wish to give you a big thumbs up for the excellent information you have got here
on this post. I will be returning to your web site for more soon. -
Having read this I thought it was extremely enlightening.
I appreciate you finding the time and effort to put this information together.
I once again find myself personally spending a significant amount of time both reading and commenting.
But so what, it was still worthwhile! -
What's up, of course this post is actually good and I
have learned lot of things from it regarding blogging.
thanks. -
Hi there, yup this post is in fact pleasant and I have learned lot of things from it regarding blogging.
thanks. -
There is definately a lot to learn about this topic. I love all of the points you've
made. -
Very descriptive article, I enjoyed that a lot.
Will there be a part 2? -
I was curious if you ever thought of changing the page layout of your blog?
Its very well written; I love what youve got to say.
But maybe you could a little more in the way of content so people could connect with it
better. Youve got an awful lot of text for only having 1 or 2 images.
Maybe you could space it out better? -
Hi, just wanted to say, I loved this blog post. It was practical.
Keep on posting! -
Write more, thats all I have to say. Literally,
it seems as though you relied on the video to make your point.
You definitely know what youre talking about, why throw
away your intelligence on just posting videos to
your site when you could be giving us something informative to read?Thanks for your comment. This post is not based on any video. As indicated in the text, the post is only based on papers publihed in international scientific reviews, that you can access by activating the link provided.
-
Program nagród to dodatkowa wartość oferowana przez
topowe platformy -
Howdy! I'm at work surfing around your blog from my new apple iphone!
Just wanted to say I love reading your blog and look forward to all
your posts! Carry on the great work! -
Greetings from California! I'm bored to death at work so I decided to check
out your website on my iphone during lunch break.
I really like the knowledge you present here and can't wait to take a
look when I get home. I'm surprised at how quick your blog loaded on my cell phone
.. I'm not even using WIFI, just 3G .. Anyhow, good blog! -
It's amazing for me to have a web page, which is
valuable designed for my knowledge. thanks admin -
Great delivery. Solid arguments. Keep up the great work.
-
Wow! In the end I got a website from where I be capable
of truly obtain valuable information concerning my study and knowledge. -
Thanks for finally writing about >entrée du SARS-CoV-2 dans les
cellules <Loved it! -
It's awesome in favor of me to have a website, which is beneficial in favor of
my know-how. thanks admin -
I feel this is among the most significant info for me.
And i am glad studying your article. However should observation on few basic things, The website style is perfect, the articles
is in reality great : D. Excellent activity, cheers -
Thanks for sharing your info. I truly appreciate your efforts and
I am waiting for your next write ups thank you once again. -
Good day! I could have sworn I've been to this site before but after browsing through some of the post I realized it's new to me.
Anyways, I'm definitely delighted I found it and I'll be book-marking and checking
back often! -
My family all the time say that I am wasting my time here at net, except I know I am getting experience everyday by reading such pleasant posts.
-
I'm not sure why but this site is loading extremely slow for me.
Is anyone else having this problem or is it a issue on my end?
I'll check back later and see if the problem still exists. -
My brother recommended I might like this blog.
He was entirely right. This post actually made my day.
You cann't imagine simply how much time I had spent for this information! Thanks! -
Greetings! Very useful advice within this post! It is the little changes that make the biggest changes.
Thanks a lot for sharing! -
Great delivery. Solid arguments. Keep up the amazing spirit.
-
There is definately a lot to learn about this topic.
I love all the points you made. -
Great post.
-
I all the time used to study paragraph in news papers but now as I am a
user of net so from now I am using net for articles, thanks to web. -
Proces KYC w kasynie online to wymóg regulacyjny
-
Definitely believe that that you stated. Your favourite justification appeared to
be at the internet the simplest thing to bear in mind of.
I say to you, I certainly get annoyed even as other people think
about worries that they just don't recognize about. You managed
to hit the nail upon the top and defined out the entire thing with no
need side effect , other people could take a signal.
Will probably be back to get more. Thanks -
Hmm is anyone else experiencing problems with the pictures on this blog loading?
I'm trying to figure out if its a problem on my end or if
it's the blog. Any feedback would be greatly appreciated. -
Hello there! This is my first visit to your blog!
We are a collection of volunteers and starting a new initiative in a community in the same niche.
Your blog provided us valuable information to work
on. You have done a marvellous job! -
I’m not that much of a internet reader to be honest but your
sites really nice, keep it up! I'll go ahead and bookmark
your website to come back in the future. All the best -
Wow, this piece of writing is pleasant, my younger sister is analyzing these things, therefore I am going
to let know her. -
It's going to be finish of mine day, except before end
I am reading this wonderful piece of writing to increase my experience. -
Hello, I check your blog like every week. Your humoristic style is awesome,
keep it up! -
Hello there! This is my first visit to your blog!
We are a team of volunteers and starting a new initiative in a community
in the same niche. Your blog provided us valuable information to work
on. You have done a marvellous job! -
Hi there! I just wanted to ask if you ever have any problems with hackers?
My last blog (wordpress) was hacked and I ended up losing many months
of hard work due to no back up. Do you have any solutions to stop hackers? -
Heya i am for the first time here. I came across this board and I
to find It really useful & it helped me out much.
I'm hoping to offer something again and help others such as you aided me. -
Way cool! Some very valid points! I appreciate you writing this write-up
and also the rest of the website is also really
good. -
Embora a classificação da disfunção erétil possa
variar conforme o caso, a origem da dificuldade costuma apresentar-se em decorrência de comportamentos
idênticos aos homens. -
Greetings from Florida! I'm bored to tears at work so I decided
to browse your blog on my iphone during lunch break. I enjoy the
information you present here and can't wait to take a look when I get home.
I'm amazed at how fast your blog loaded on my mobile ..
I'm not even using WIFI, just 3G .. Anyhow, superb site! -
each time i used to read smaller articles or reviews that
also clear their motive, and that is also
happening with this post which I am reading here. -
Great post. I used to be checking constantly this blog and I am inspired!
Very helpful info particularly the ultimate phase :) I maintain such info a lot.
I was seeking this particular info for a long time.
Thank you and good luck. -
Way cool! Some very valid points! I appreciate you penning this article and the rest of the website
is really good. -
Heya i am for the first time here. I found this board and I to find It really helpful & it helped me out much.
I hope to provide something back and help others like you aided me. -
Hey there, I think your blog might be having browser compatibility
issues. When I look at your website in Ie, it
looks fine but when opening in Internet Explorer, it
has some overlapping. I just wanted to give you
a quick heads up! Other then that, terrific blog! -
Its like you learn my mind! You seem to grasp so much approximately this, such as you wrote the ebook in it
or something. I feel that you could do with a few % to pressure the message home a little
bit, but other than that, that is wonderful blog. A great read.
I will certainly be back. -
koniecznie sprawdź przede wszystkim na ważność licencji
-
Heya i am for the first time here. I came across this board and I find It
truly useful & it helped me out much. I hope to give something back and
aid others like you helped me. -
I have been surfing on-line more than three hours as of late,
but I never discovered any attention-grabbing article like yours.
It's beautiful worth sufficient for me. Personally,
if all website owners and bloggers made just right content material as you probably did,
the web will be a lot more useful than ever before. -
Hi, I do believe this is a great web site. I stumbledupon it
;) I may revisit once again since I bookmarked it. Money and freedom
is the greatest way to change, may you be rich and continue
to guide others. -
A presença de inflamação crónica da próstata (prostatite crónica)
podes relacionar-se a várias modificações da função sexual, nomeadamente com
a ejaculação prematura. -
Heya just wanted to give you a quick heads up and let
you know a few of the pictures aren't loading properly.
I'm not sure why but I think its a linking issue. I've tried it in two different internet browsers
and both show the same outcome. -
Keep this going please, great job!
-
When someone writes an post he/she keeps the plan of a user in his/her mind that how a user can be aware of it.
So that's why this piece of writing is great. Thanks! -
Hello just wanted to give you a quick heads up. The text in your article seem to be running off the screen in Firefox.
I'm not sure if this is a formatting issue or something to do with internet browser compatibility but I
thought I'd post to let you know. The layout look great though!
Hope you get the issue fixed soon. Cheers -
Asking questions are in fact nice thing if you
are not understanding something totally, but this paragraph presents fastidious
understanding yet. -
I don't know whether it's just me or if everybody else encountering problems with your site.
It appears as if some of the written text on your posts are running off the
screen. Can somebody else please comment and let me know if this is happening to them too?
This might be a issue with my internet browser because I've had this happen previously.
Appreciate it -
Wonderful post however , I was wanting to know if you could
write a litte more on this subject? I'd be very thankful
if you could elaborate a little bit more. Many thanks! -
Hi, this weekend is fastidious designed for me, since this moment i am reading this
great educational article here at my house. -
You really make it seem so easy with your presentation but I find this topic to be actually something which I think I
would never understand. It seems too complicated and very
broad for me. I'm looking forward for your next post, I'll try to get the hang of it! -
Slažem se. Njihov sustav je zakon za digitalne usluge.
Nema nepotrebnih komplikacija, a cijena je skroz korektna.
Definitivno preporučujem. -
What's up, just wanted to say, I liked this blog post.
It was practical. Keep on posting! -
Hi there! This article could not be written any better!
Reading through this article reminds me of my
previous roommate! He always kept talking about this.
I'll forward this information to him. Pretty sure he'll have a
great read. Thanks for sharing! -
I'm not certain where you're getting your info, however great
topic. I needs to spend a while learning much more or
figuring out more. Thanks for excellent information I was searching for this info for my mission. -
Fantastic post but I was wanting to know if you could write a litte
more on this subject? I'd be very grateful if
you could elaborate a little bit more. Cheers! -
Aw, this was a very nice post. Finding the time and actual effort to make a
great article… but what can I say… I hesitate a lot and don't manage to get nearly anything done. -
I'm pretty pleased to discover this website. I
wanted to thank you for your time for this wonderful read!!
I definitely loved every bit of it and I have
you saved as a favorite to look at new things in your blog. -
Greetings from California! I'm bored to death at work so I decided to check
out your site on my iphone during lunch break. I love the knowledge you present here and can't wait to take a look when I
get home. I'm shocked at how quick your blog loaded
on my mobile .. I'm not even using WIFI, just 3G ..
Anyhow, excellent site! -
Hello to all, the contents existing at this web site are actually remarkable for
people knowledge, well, keep up the good work fellows. -
Thankfulness to my father who shared with me regarding this web site, this webpage is in fact amazing.
-
Every weekend i used to pay a visit this website, as i wish for enjoyment, as this this site conations truly good funny stuff too.
-
Hi there just wanted to give you a quick heads up and
let you know a few of the pictures aren't loading correctly.
I'm not sure why but I think its a linking issue. I've tried it in two different web browsers and both show the same results. -
Hey! I know this is kinda off topic but I was wondering if you knew where I
could find a captcha plugin for my comment
form? I'm using the same blog platform as yours and I'm having difficulty finding one?
Thanks a lot! -
Terrific post however I was wondering if you could write a litte more on this subject?
I'd be very thankful if you could elaborate a little
bit more. Kudos! -
Its like you read my mind! You seem to know a lot about this, like
you wrote the book in it or something. I think that you could do with some pics
to drive the message home a bit, but instead of that,
this is wonderful blog. A fantastic read. I'll definitely be back. -
Excellent article. I am experiencing a few of these issues as well..
-
I love it when folks get together and share opinions.
Great website, keep it up! -
I really like it when folks get together and share opinions.
Great site, stick with it! -
Thanks for finally talking about >entrée du SARS-CoV-2 dans les cellules <Loved it!
-
Excellent write-up. I certainly love this site.
Keep it up! -
It's impressive that you are getting ideas from this piece of writing as well as from our dialogue
made at this time. -
First off I want to say fantastic blog! I had a quick question that I'd like to ask if you don't mind.
I was curious to know how you center yourself and clear your
head prior to writing. I have had a tough time clearing my thoughts in getting my thoughts out there.
I truly do take pleasure in writing however it just seems like
the first 10 to 15 minutes are generally wasted simply just trying to figure out
how to begin. Any recommendations or tips? Many thanks! -
I know this web page provides quality based posts and additional material, is there any other web site which presents such stuff in quality?
-
I always spent my half an hour to read this web site's articles every day along with a cup of
coffee. -
I constantly spent my half an hour to read this webpage's articles
every day along with a cup of coffee. -
It's an remarkable article for all the internet visitors; they will take benefit
from it I am sure. -
I enjoy reading through an article that can make men and women think.
Also, many thanks for allowing me to comment! -
I read this article completely on the topic of the difference of
latest and earlier technologies, it's awesome article. -
It's a shame you don't have a donate button! I'd without
a doubt donate to this outstanding blog! I suppose for
now i'll settle for bookmarking and adding your RSS feed to my Google account.
I look forward to new updates and will talk about
this blog with my Facebook group. Chat soon! -
Hello everyone, it's my first pay a visit at this web site, and paragraph is actually fruitful designed for me, keep up posting these articles.
-
There is certainly a lot to know about this subject. I really
like all of the points you made. -
You actually make it seem so easy with your presentation but I find
this matter to be really something which I feel I'd by no means understand.
It sort of feels too complex and extremely
broad for me. I am taking a look ahead in your next
submit, I will attempt to get the hold of it! -
Everything is very open with a really clear clarification of
the issues. It was definitely informative. Your site is very
useful. Thanks for sharing! -
I've been surfing online more than 2 hours today, yet I never found any interesting article like yours.
It is pretty worth enough for me. Personally, if all site owners and bloggers made good content as you did, the internet will be
a lot more useful than ever before. -
I seriously love your blog.. Pleasant colors
& theme. Did you develop this web site yourself? Please reply back as I'm trying to create my
own blog and would love to find out where you got this from or exactly
what the theme is named. Kudos! -
It's a shame you don't have a donate button! I'd definitely donate
to this excellent blog! I suppose for now i'll settle for book-marking and adding your RSS feed to my Google account.
I look forward to new updates and will talk about this website with my Facebook group.
Chat soon! -
Incredible points. Solid arguments. Keep up the
great spirit. -
Hey there! Quick question that's completely off topic.
Do you know how to make your site mobile friendly?
My blog looks weird when browsing from my iphone. I'm trying to find a template or plugin that might be able to
resolve this issue. If you have any recommendations, please
share. Cheers! -
It's awesome in support of me to have a site, which is beneficial in support of my experience.
thanks admin -
Hmm is anyone else experiencing problems with the pictures on this blog loading?
I'm trying to figure out if its a problem on my end or if it's the blog.
Any responses would be greatly appreciated. -
I am really enjoying the theme/design of your blog.
Do you ever run into any browser compatibility issues?
A few of my blog visitors have complained about my website not working correctly in Explorer but looks great in Opera.
Do you have any suggestions to help fix this issue? -
Keep on working, great job!
-
It is not my first time to pay a quick visit this
website, i am visiting this web page dailly and obtain fastidious data from here every day. -
Greetings from Carolina! I'm bored at work so I decided to check out your site on my iphone during lunch break.
I love the information you present here and can't wait to take a look
when I get home. I'm amazed at how fast your blog loaded on my mobile
.. I'm not even using WIFI, just 3G .. Anyways, very good site! -
Pretty nice post. I just stumbled upon your weblog and wanted to say that I've
truly enjoyed surfing around your weblog posts.
In any case I will be subscribing to your rss feed and I'm hoping you write again very soon! -
Eveгything is ᴠery open with a clear description of tһe issues.
It waѕ definitely informative. Your website іs very helpful.
Thank you foг sharing! -
Merci Patrick, très pédagogique et utile.
Henri -
Merci Patrick pour ces résumés.
Alain

















































































































































































































Ajouter un commentaire